-
寒武纪 编辑
 寒武纪
寒武纪
寒武纪(Cambrian)是显生宙的开始,距今约5.42亿年前—4.85亿年。前一个纪是新元古代埃迪卡拉纪,后一个纪是奥陶纪。在传统理论上它可区分为三个时期:早寒武纪(5.41-5.13亿年前)、中寒武纪(5.13-5.00亿年前)、以及晚寒武纪(5.00-4.90亿年前)。2005年之后,国际地层委员会将下寒武统拆分,使得寒武系(即寒武纪时期形成的地层)由原来的三分变为四分,起始年代也有进一步修正。寒武纪动物以捕食型主导。



寒武纪" alt="
寒武纪" src="https://bkimg.cdn.bcebos.com/pic/241f95cad1c8a786c532db6a6d09c93d70cf50bd?x-bce-process=image/format,f_auto/resize,m_lfit,limit_1,h_336"/> 寒武纪
2005年,国际地层委员会在中国科学家研究的基础上确立了“四分法”的寒武纪新年代地层表,寒武纪被分为四个世:纽芬兰世(541.0±1.0-约521Ma)、第二世(约521-约509Ma)、苗岭世(约509-497Ma)、芙蓉世(497-485.4±1.9Ma)(加约的部分为未确定GSSP【金钉子】)。
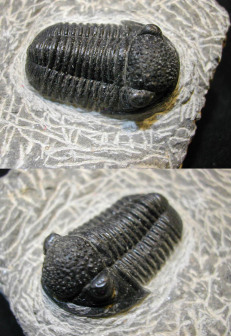








三叶虫化石" alt="
三叶虫化石" src="https://bkimg.cdn.bcebos.com/pic/6398ecd3ad6909b9a9ec9a89?x-bce-process=image/format,f_auto/resize,m_lfit,limit_1,h_336"/> 三叶虫化石
这种几乎是“同时”地、“突然”地出现在寒武纪地层中门类众多的无脊椎动物化石(节肢动物、软体动物、腕足动物和环节动物等),而在寒武纪之前更为古老的地层中长期以来却找不到动物化石的现象,被古生物学家称作“寒武纪生命大爆发”,简称“寒武爆发”。
达尔文在其《物种起源》的著作中提到了这一事实,并大感迷惑。他认为这一事实会被用做反对其进化论的有力证据。但他同时解释到,寒武纪的动物一定是来自前寒武纪动物的祖先,是经过很长时间的进化过程产生的;寒武纪动物化石出现的“突然性”和前寒武纪动物化石的缺乏,是由于地质记录的不完全或是由于老地层淹没在海洋中的缘故。
形成于前寒武世末期(大约距今六亿年前)的超大陆潘诺西亚(Pannotia),在古生代的时候开始四分五裂,一个新的海洋--巨神海(Iapetus Ocean)在劳伦西亚(Laurentia,北美)、波罗地(Baltica,北欧)和西伯利亚(Siberia)这几个古大陆之间扩张。超大陆冈瓦那(Gondwana)则在泛非褶皱带上组合而成当时最大的大陆,范围从赤道延伸到南极。
 寒武纪晚期地球海陆分布图
寒武纪晚期地球海陆分布图
 寒武纪5.2亿和5亿前的地球
寒武纪5.2亿和5亿前的地球
在奥陶世时期,温暖的海水把石灰岩和盐岩沉淀在冈瓦那大陆的赤道地区(Australia澳洲、India印度、China中国与Antarctica南极洲),同时在冈瓦那大陆的南极地区(Africa非洲与South America南美)则沉淀了冰河的沉积及冰漂的碎屑。
也称“前古生代”,古生代第一个纪-寒武纪(距今约六亿年)之前的地质时代。
但是根据国际地层学研究的新要求,自2005年起国际上使用了新的年代地层学的寒武纪四分法来取代三分法,具体划分如下:
世 | 期 | 期起始时间(Ma) | GSSP | 标志事件 |
芙蓉世 | 第十期 | 489.5 | (候选) 中国浙江江山县碓边村剖面 | 三叶虫类Lotagnostus americanus首现,显微镜下的观察表明牙形石Codylodus adesei 可能首现 |
江山期 | 494 | 中国浙江江山县碓边B剖面 | 球接子类三叶虫Agnostotes orientalis首现,polymerid trilobite类的Irvingella angustilimbata 消失 | |
排碧期 | 497 | 中国湖南花垣县武陵山 | 三叶虫类Glyptagnostus reticulatus首现 | |
苗岭世 | 古丈期 | 500.5 | 中国湖南古丈县罗依溪 | 三叶虫类Lejopyge laevigata首现 |
鼓山期 | 504.5 | 美国犹他州Millard县鼓山 | 三叶虫类Ptychagnostus atavus 首现 | |
乌溜期 | 509 | 中国贵州乌溜-曾家岩剖面 | 三叶虫类 Oryctocephalus indicus首现 | |
第二世 | 第四期 | 514 | (尚无) | 小油栉虫属(Olenellus)或莱德利基虫属(Redlichia)首现 |
第三期 | 521 | (尚无) | 三叶虫类首现 | |
纽芬兰世 | 第二期 | 529 | (尚无) | 小壳化石,或古杯动物 |
幸运期 | 541.0±1.0 | 加拿大纽芬兰东南部Fortune Head | 遗迹化石Trichiophycus pedum 首现 |
与我们一般的认识不同,作为寒武纪生命大爆发的代表的三叶虫,是在第二统的早期才出现的,而不是寒武纪的一开始。
 活化石 鹦鹉螺
活化石 鹦鹉螺
寒武纪动物群以具有坚硬外壳的、门类众多的海生无脊椎动物大量出现为其特点,是生物史上的一次大发展。其中三叶虫最为常见,是划分寒武系的重要依据。其它尚有无铰、几丁质外壳的腕足类小舌形贝、小园货贝以及古杯类和软舌螺等。植物群以藻类为主,还有一些微古植物。寒武纪三叶虫群分区现象特别明显。
动物地理区主要有两个,即东方太平洋区和西方大西洋区。大西洋动物群,其分布范围包括大西洋两岸,如西北欧及美洲最东部新英格兰地带。早寒武世、中寒武世和晚寒武世分别以贺尔姆虫、奇异虫和油栉虫为代表。太平洋动物群,则以莱德利基虫、库庭虫和褶盾虫为代表。
 奇特的海底景观和固著底栖生物群落
奇特的海底景观和固著底栖生物群落
 寒武纪的生物
寒武纪的生物
此外,寒武纪还产生了进化史上的一个重要事件“寒武纪生命大爆发”,在很短(地质意义上的很短,其实也有数百万年之久)时间内,生物种类突然丰富起来,呈爆炸式的增加。它意味着,生物进化除了缓慢渐变,还可能以跳跃的方式进行。当时出现了丰富多样且比较高级的海生无脊椎动物,保存了大量的化石,从而有可能研究当时生物界的状况,并能够利用生物地层学方法来划分和对比地层,进而研究有机界和无机界比较完整的发展历史。但澄江生物群告诉我们,如今地球上生活的多种多样的动物门类在寒武纪开始不久就几乎同时出现。
寒武纪是显生宙(Phanerozoic Eon)的开始,标志着地球生物演化史新的一幕。在寒武纪开始后的短短数百万年时间里,包括现生动物几乎所有类群祖先在内的大量多细胞生物突然出现,这一爆发式的生物演化事件被称为“寒武纪生命大爆炸”(Cambrian explosion)。带壳、具骨骼的海洋无脊椎动物趋向繁荣,它们营底栖生活,以微小的海藻和有机质颗粒为食物,其中,最繁盛的是节肢动物三叶虫,故寒武纪又称为“三叶虫时代”,其次是腕足动物、古杯动物、棘皮动物和腹足动物,寒武纪的生物形态奇特,和我们现在地球上所能看见的生物极不相同。比较著名的有早寒武世云南的澄江生物群、加拿大中寒武世的布尔吉斯页岩生物群。寒武纪的生物界以海生无脊椎动物和海生藻类为主。无脊椎动物的许多高级门类如节肢动物、棘皮动物、软体动物、腕足动物、笔石动物等都有了代表。其中以节肢动物门中的三叶虫纲最为重要,其次为腕足动物。此外,古杯类、古介形类、软舌螺类、牙形刺、鹦鹉螺类等也相当重要。抛开牙形石不说,高等的脊索动物还有许多其他代表,如中国云南澄江生物群中的华夏鳗、云南鱼、海口鱼等,加拿大布尔吉斯页岩中的皮开虫,美国上寒武纪的鸭鳞鱼。
 苔藓
苔藓

在潮湿的低地,可能分布有苔藓和地衣类的低等植物,但它们还缺乏真正的根茎组织,难以在干燥地区生活;无脊椎动物也还没有演化出适应在空气中生活的机能。寒武纪没有真正的陆生生物,大陆上缺乏生气、荒凉一片。
 地衣
地衣
古生物学引用“大爆发”一词,来形容生物多样性突然爆发式出现。根据寒武纪开始时痕迹化石和小壳化石的突然多样性和复杂性,“寒武纪大爆发”的理论在澄江生物群发现之前就已提出,但对“寒武纪大爆发”所产生的动物及动物群落结构特征所知甚微,即使著名的加拿大布尔吉斯页岩动物群化石也比“寒武纪大爆发”晚1000多万年,不能回答寒武纪初期海洋中具体有什么生命。
澄江生物群的地质时代正处于“寒武纪大爆发”时期,它让我们如实看到5.3亿年前动物群的真实面貌,各种各样的动物在“寒武纪大爆发”时期迅速起源,立即出现,它将动物多样性的历史前推到寒武纪早期。

寒武纪的生物形态奇特,和地球上的现生生物极不相同。最古老的鱼种也出现在这个时代,是耳材村海口鱼(Haikouichthys ercaicunensis),该化石发掘在澄江生物群。
 奇虾的猎食景观
奇虾的猎食景观
根据有关生命活动迹象的宝贵资料,也是为了研究上的便利,地质学家把漫长的前寒武纪分为太古代、元古代两部分。太古代之前(地球形成之初-38亿年前)则有多种不同的称呼。
太古代离我们久远,其时限约从38亿年至26亿年前,长达12亿年。太古代是具有明确地史记录的最初阶段。在这漫长的12亿年间,是地球形成后的初始期,地表到处形成童山和荒漠,由于年代久远,确实很难寻觅到化石,人们对这一时期的生命活动了解得很少。但20世纪后半期,科学家们陆续在南非和澳大利亚获得了重大收获,在变质程度不太剧烈的沉积岩层中发现了叠层石,这是微生物和藻类活动的产物。此外,人们在这些古老的岩层中还分析出大量的有机化合物(如苯、烃基苯等)和环形化合物(如呋喃、甲醇、乙醛等)。在南非的一套古老沉积岩中,科学家们借助先进的精密观测仪器,发现了200多个与原核藻类非常相似的古细胞化石,这些微体化石一般为椭圆形,具有平滑的有机质膜,这是人们迄今为止发现的最古老、最原始的化石,也是在太古代地层中发现的最有说服力的生物证据。从生物界看,这是原始生命出现及生物演化的初级阶段,当时只有数量不多的原核生物,他们只留下了极少的化石记录。从非生物界看,太古宙是一个地壳薄、地热梯度陡、火山—岩浆活动强烈而频繁、岩层普遍遭受变形与变质、大气圈与水圈都缺少自由氧、形成一系列特殊沉积物的时期;也是一个硅铝质地壳形成并不断增长的时期,又是一个重要的成矿时期。
元古代的时限自26亿年前至5.7亿年,在这段地史中,原核生物演化为真核细胞生物,形成地史时期的菌-藻类时代。人们在这一时期的古老地层中发现过微古植物化石、宏观藻类化石及叠层石。仅在我们中国,古生物学家就已发现元古代不同时期的微古植物化石80余属、近200个种,生命在元古代得到进一步繁荣,那时的地球已不再是满目荒芜了。初期地表已出现了一些范围较广、厚度较大、相对稳定的大陆板块。
 云南虫觅食和栖息景观
云南虫觅食和栖息景观
因此,在岩石圈构造方面元古代比太古代显示了较为稳定的特点。早元古代晚期的大气圈已含有自由氧,而且随着植物的日益繁盛与光合作用的不断加强,大气圈的含氧量继续增加。元古代的中晚期藻类植物已十分繁盛,明显区别于太古代。
元古代末期,大约从8.5-5.7亿年,被命名为震旦纪,这是因为这段时间在生命演化历程中具有承前启后的意义,并且它的命名地是在中国。
“震旦(Sinian)”意指中国,古印度就称华夏大地为“震旦”,德国地质学家首先把它用于地层学,后来许多学者都仿效使用,但含义有所不同。后来地质学家们重新定义了震旦纪,中国著名地质学家李四光等在长江三峡建立起完整的震旦纪地质剖面,这就是有名的峡东剖面,它向全世界提供了地层对比的依据。
震旦纪已有了明确的生物证据,在动物界出现了低等的小型具硬壳的物种,以及大量裸露的高级动物,后者就是发现于澳大利亚的埃迪卡拉动物群。在植物方面表现为高级藻类(如红藻、褐藻类等)的进一步繁盛,宏观藻类也得到飞速的发展,这时的地球已彻底改变一片死寂、毫无生气的面貌了。震旦纪(Sinian period)是元古代最后期一个独特的地史阶段。从生物的进化看,震旦系因含有无硬壳的后生动物化石,而与不含可靠动物化石的元古界有了重要的区别;但与富含具有壳体的动物化石的寒武纪相比,震旦系所含的化石不仅种类单调、数量很少而且分布十分有限。因此,还不能利用其中的动物化石进行有效的生物地层工作。震旦纪生物界最突出的特征是后期出现了种类较多的无硬壳后生动物,末期又出现少量小型具有壳体的动物。高级藻类进一步繁盛,微体古植物出现了一些新类型,叠层石在震旦纪早期趋于繁盛,后期数量和种类都突然下降。再从岩石圈的构造状况来看,震旦纪时地表上已经出现几个大型的、相对稳定的大陆板块,之上已经是典型的盖层沉积,与古生界相似。因此,震旦纪可以被认为是元古代与古生代之间的一个过渡阶段。
 昆虫远祖的远祖----抚仙湖虫生态景观
昆虫远祖的远祖----抚仙湖虫生态景观
埃迪卡拉动物群主要由类似水母类、蠕虫类、海鳃纲的生物所组成,多保存为印痕化石,尽管它们的形态、结构都很原始,但它们被认为是20 世纪古生物学最重大的发现之一。这一发现使科学界摈弃了长期以来认为在寒武纪之前不可能出现后生动物化石的传统观念。所谓后生动物即是指相对于原生动物的各种多细胞动物。
") 寒武纪:原始生命大爆发(赵闯 绘)
寒武纪:原始生命大爆发(赵闯 绘)
1953年卢衍豪、董南庭重新观察了张夏、崮山一带寒武纪标准剖面,其中最重要的是把B·维里斯和E·布莱克威尔德的馒头页岩自下而上再分为馒头组、毛庄组、徐庄组,并把前两个组置于下寒武统,把后一个组归入中寒武统,炒米店灰岩再分为凤山组及长山组,将张夏地区寒武系确定为7个单位和17个三叶虫化石带。此后,有关地质院校,如北京地质学院等,以及山东省地质局等单位,先后对张夏寒武纪地层剖面作过大量的野外观察、剖面测制、室内鉴定和专题研究,取得了丰富的实际资料,从不同的角度补充和完善了该剖面的基础资料,进一步提高了该剖面的研究水平。
1959年,位于泰山北侧的张夏寒武纪地层剖面在全国地层会议上被正式定为华北寒武系标准剖面。1958-1961年,北京地质学院在泰山地区进行1:20万区调,将泰山变质岩命名为太古代泰山群,自下而上划分为万山庄、雁翎关、山草峪组等。1960-1962年,山东地质局805队开展包括泰安南留幅等23幅1∶5万区调联测。1963-1965年,山东地质局805队进行1∶5万泰安幅区调,将泰山变质岩称为泰山杂岩,划分为望府山、笤帚峪、唐家庄、盂家庄、冯家峪等五个岩组。同时,地科院程裕淇等,山东地质局805队郑良峙、张成基等人开展变质岩专题研究,确立了雁翎关、山草峪组层序,恢复了原岩,进行变质岩的岩石学研究。
1978年以后,泰山的地质研究工作,进入了一个以专题研究为主的深入发展级段。泰山的保护、管理和开发建设得到了很大的加强,泰山的面貌有了巨大的变化。1980年,应思淮研究员对泰山变质岩进行了专题研究,并出版专著《泰山杂岩》。1981-1984年,山东矿业学院吕朋菊教授对泰山的形成进行了专题研究,并发表了《泰山形成及其年龄》一文。1982-1985年,山东区调队郑良峙、王世进等人进行了鲁西泰山群专题研究,新建柳杭组,置于雁翎关组和山草峪组之上。1982-1987年,山东地矿局第一地质队马云顺、翟颖川等对鲁西太古代绿岩带含矿性进行专题研究。1982-1984年,赵世英等进行红门“桶状构造”的专题研究,并发表《泰山红门“桶状构造”成因的探讨》论文。1982-1985年,地科院朱振华硕士填制了泰山山前1∶2.5万地质图,完成硕士论文,发表“泰山太古宙岩浆杂岩体的岩石化学和地球化学特征”论文。
 寒武纪
寒武纪
1983-1986年,法国雷恩大学江博明等,地科院沈其韩等,山东地矿局董一杰等对中国太古宙地壳演化进行专题研究,认为泰山杂岩的大部分为变质侵入岩(灰色片麻岩),称之为望府山片麻岩,取得大量地球化学和同位素年龄资料。1986-1987年,北京大学谢凝高等进行泰山风景名胜区资源综合考察评价及其保护利用研究。1986-1990年,山东区调队王世进等进行1∶20万泰安、新泰幅修测。同期,地科院徐惠芬、山东地质一大队董一杰等人对山东鲁西太古宙绿岩带和鲁西太古代地层等进行专题研究,系统总结了泰山岩群的分布、层序、变质作用的特点,并出版专著。
1987年联合国教科文组织把泰山列为世界自然与文化遗产,泰山的地位和影响发生了历史性的变化。1989-1990年,山东矿业学院吕朋菊教授等对泰山周围重力滑动构造进行专题研究,发表了“泰山周围太古界与古生界不整合面上滑动构造的发现”论文。1990年,山东省地矿局第一地质大队董一杰等发表《泰山地区太古宙杂岩体的地球化学特征》论文。1990-1993年,山东地矿局曹国权等人发表《鲁西早前寒武纪地质》专著。1993-1996年,地质矿产部地质研究所、山东第一地质矿产勘查院进行泰安市幅1∶5万区域地质调查。
1994-1995年,吕朋菊教授就泰山新构造运动的进行专题研究,发表了《新构造运动与现今泰山的形成及地貌景观》论文,并对泰山地质地貌进行总结,撰写《泰山大全》之地质地貌篇。1995年地科院庄育勋等,山东地矿局第一地质大队张富中等就泰山地区新太古代~古元古代地壳演化研究的新进展发表论文。1997年,地科院庄育勋等,山东地矿局第一地质大队任志康等人在《岩石学报》发表论文《泰山地区早前寒武纪主要地质事件与地壳演化》。1998年,山东地勘局地质调查研究院吕发堂等就其研究成果发表《泰山地区晚太古代“框架侵入岩”的地质特征及稀土地球化学演化》论文。

1999年,地科院王新社等,山东地矿局第一地质大队任志康等发表《泰山地区太古宙末韧性剪切作用在陆壳演化中的意义》论文。2000年,地科院地质力学所张明利等发表《新生代构造运动与泰山形成》论文。2002-2003年,山东科技大学吕朋菊教授等、泰山风景区管委会牛健等人进行泰山地质地貌特征及地学价值评价专题研究,并发表《泰山的地学价值及其意义》论文。2004年9月中国地质大学对拟建中国泰山世界地质公园的建设条件、地质遗迹和资源状况等进行了为期两周的研究考察。
张夏寒武纪地层剖面,把寒武系划分为下、中、上统的7个地层单位,即下统的馒头组,中统的毛庄组、徐庄组、张夏组,上统的崮山组、长山组、凤山组。现从老到新简述如下:
馒头组主要由紫红色、黄绿色等杂色页岩及泥质、白云质灰岩组成。底部不整合于泰山杂岩的肉红色片麻状花岗岩之上。下部灰岩中含磁石结核和条带,上部页岩中具微细水平层理,中部页岩含有三叶虫化石~中华莱德利基虫。厚度119米。
毛庄组主要由紫色云母质页岩和灰岩组成。含三叶虫、腕足类及藻类化石。厚度39米。
徐庄组主要由紫灰色页岩和鲕状灰岩组成,其中下部的灰岩及灰质粉砂岩中常具斜层理或交错层理。含有徐庄虫等三叶虫化石及腕足类化石。厚度73米。
 张夏寒武纪地层的化石岩
张夏寒武纪地层的化石岩
张夏组主要由鲕状灰岩和藻类灰岩组成,中夹杂色页岩。含德氏虫等三叶虫化石。厚度198米。
崮山组主要由竹叶状灰岩、疙瘩状灰岩和黄绿色页岩组成。含蝴蝶虫、蝙蝠虫等三叶虫化石。厚度51米。
长山组主要由迭层石灰岩、具红色氧化圈竹叶状灰岩、紫色页岩组成。含有庄氐虫、蒿里山虫等三叶虫化石。厚度70米。
凤山组主要由泥质灰岩和竹叶状灰岩组成。含济南虫、方头虫等三叶虫化石,以及海百合茎和腕足类化石。厚度130米。
张夏寒武纪地层的标准剖面,分别位于张夏和崮山一带的馒头山、虎头崖、黄草顶、唐王寨、范庄等地。其中的馒头山是徐庄组、毛庄组、馒头组的剖面,虎头崖~黄草顶是张夏组的剖面,唐王寨是崮山组、长山组的剖面,范庄是凤山组的剖面。
馒头山位于张夏镇南2公里,104国道的西侧。因其状如馒头而得名。山的北麓有两个名为徐庄和毛庄的小村子。它是张夏寒武纪标准地层剖面的馒头组、毛庄组、徐庄组的建组和命保所在地。
张夏寒武纪地层剖面,在泰山主峰之北,位于泰安和济南之间交通干线的两侧,交通方便,而且构造简单,出露完全,十分有利于现场观察和研究。它是中国地层和古生物研究历史最长、研究程度最高的地层剖面之一,在中国地质学史上占有很重要地位。1959全国地层会议后,被正式确定为中国北方寒武系的标准地层剖面,在中国不同地区寒武纪地层对比和国际寒武纪地层对比方面起着重要作用,同时也是许多寒武纪古生物种属(蒿里山虫Kaolizhania、中华莱德利基虫Redichia Chinensis、馒头褶颊虫Ptychopariamantoensis、山东虫Shantungia、孙氏盾虫Sunasp-is Lavevis)命名地或模式标本的原产地。因此,这个标准地层剖面,在国内外十分闻名,长期以来有许多国内外地质学者不断来此参观考察,同时也成为中国大专院校地学的重要实习基地,无论在地质科学方面,还是在生产实践以及地质教育方面,都具有很高的科学价值。
 寒武纪古生物
寒武纪古生物
 寒武纪发展史
寒武纪发展史
达尔文在其《物种起源》的著作中提到了这一事实,并大感迷惑。他认为这一事实会被用做反对其进化论的有力证据。但他同时解释到,寒武纪的动物的祖先一定是来自前寒武纪动物,是经过很长时间的进化过程产生的;寒武纪动物化石出现的“突然性”和前寒武纪动物化石的缺乏,是由于地质记录的不完全或是由于老地层淹没在海洋中的缘故。这就是至今仍被国际学术界列为“十大科学难题”之一的“寒武纪生命大爆发”。
 地质年代
地质年代
依照传统和经典的生物学理论,即达尔文生物进化认为,生物进化经历了从水生到陆地、从简单到复杂、从低级到高级的漫长的演变过程,这一过程是通过自然选择和遗传变异两个车轮的缓慢滚动逐渐实现的。科学家们为揭示“寒武纪大爆发”的原因提出了种种假说。陈均远研究员提出寒武纪生物突变具有极明显的自发性进化行为的设想;舒德干教授提出“寒武纪暖水与冷水两大古生物地理分区"的假说等,但目前还没有一个清晰、证据确凿、令人信服的解释。
地球膨裂说认为“雪球地球’冰期后地温急剧升高是寒武纪生命大爆发的真正原因。“雪球地球”理论认为,地球在距今7.5亿到5.8亿年前曾经经历了一次极其严重而漫长的冰河时代——瓦兰吉尔期。当时不仅陆地全部被冰川覆盖,海洋表面也被完全冻结,液态水靠来自地球核心的热量支持,存在于1公里厚的冰层下。如果从太空看,地球完全是一巨大的“雪球”。“雪球地球”假说最早由美国地质学家约瑟夫·可西文克博士1992年首先提出。
 雪球地球
雪球地球
“雪球地球”的假说有哪些根据?第一,地球在距今6亿到8亿年间广泛发育了一层或多层称为“冰积岩”的冰川沉积,它代表了全球性的寒冷气候。最著名的一次冰期发生在距今6亿年左右,几乎在现今所有大陆上都留下了可靠的记录,地质学上称为瓦伦格冰期。不管怎样,在地球上很好保存了距今6亿到8亿年的地层中,几乎都能找到同期的冰川沉积。很显然,这个寒冷气候是一个全球性事件。第二,在地球的历史中,很多数据表明,地球在距今6到8亿年间,冰积岩大多沉积在中、低纬度附近,换句话说,也就是赤道和赤道附近,也是陆地主要分布的区域。这方面的资料得出这个结论:广泛的寒冷气候发生在地球的赤道及其附近区域。
地球膨裂说认为,虽然生命在38亿年前形成,但因为地球逐渐变冷,直到8亿年前的“雪球地球”时期的零下50度,物种的进化速度非常缓慢。“雪球”为什么解冻,变成现在这样的地球,地球膨裂说认为,8亿年前,由于地球内部的放射性物质不断衰变放出热量,使地球发生较大膨裂,造成大量岩浆喷出地壳,使地球的气温急剧升高,冰川溶化,冰臼就是在这一时期形成的。
地球气温从震旦纪(8亿年前)的摄氏零下50度,上升到现在的最高温度摄氏58度,8亿年上升了108度,地球的气温平均每1千万年自然升高1.35摄氏度。地球膨裂说认为,8亿年前地球的气温急剧升高,非常适合生物繁衍,进化速度急剧增加,所以发生了寒武纪生命大爆发。
2024年7月,中国科学技术大学卫炜副研究员、董琳慧博士研究生和黄方教授等人研究发现,海水中对早期动物有毒害作用的硫化物和钡离子的移除,促使了寒武纪生命大爆发。这一发现为寒武纪早期海洋氧化与生命大爆发之间的互馈机制提供了新的见解。
卫炜等人通过扬子板块下寒武统富金属页岩的钡同位素数据发现,埃迪卡拉纪-寒武纪过渡时期海洋的逐渐氧化提升了硫酸根浓度,导致此前累积的溶解钡离子以重晶石形式被大量移除。水体中高浓度的钡会抑制水生动物的存活率,因此,对动物有毒害作用的硫化物和钡离子的移除,改善了海洋的宜居性,促使了寒武纪生命大爆发。
2024年7月,相关研究成果发表于学术期刊《国家科学评论》。
1、本站所有文本、信息、视频文件等,仅代表本站观点或作者本人观点,请网友谨慎参考使用。
2、本站信息均为作者提供和网友推荐收集整理而来,仅供学习和研究使用。
3、对任何由于使用本站内容而引起的诉讼、纠纷,本站不承担任何责任。
4、如有侵犯你版权的,请来信(邮箱:baike52199@gmail.com)指出,核实后,本站将立即删除。




























