-
头索动物亚门 编辑
 头索动物亚门
头索动物亚门
头索动物栖息在热带和亚热带的浅海中,体呈鱼形。头索动物的构造虽然简单原始,但是脊索动物的三大特征都以简单的形式终生保留着.因而被称为一个典型的脊索动物的简化缩影.在动物学研究中占有重要地位。
头索动物亚门
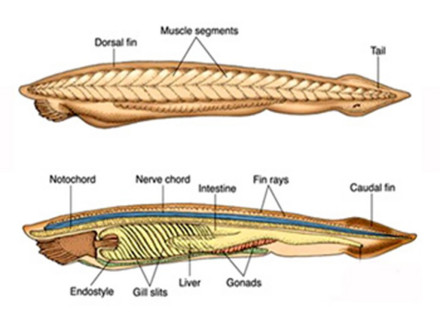
其典型代表种Branchostoma(文昌鱼);体呈鱼形,长40毫米左右,侧扁,头尾尖削,无头与躯干部之分,故名。体半透明,呈透明,呈微红色,生活于浅海疏松的沙滩里,常钻孔。以矽藻等为食。沿背侧有一纵行皮肤褶皱,称为背鳍;再后延至尾部高起,成为尾鳍,作矛形。自尾鳍腹面向前至体后约1/3处的皮肤褶襞,称为腹褶(metapleure)。左右两腹褶向后的汇合处是为腹孔,或围鳃腔孔(abdominal pore),腹鳍与尾鳍之间的小孔,为肛门。头部前端突出部位为吻。其后的腹面有一环形薄膜,成为口笠。其边缘生有36-50条触须。口笠的内腔称为前庭。
神经管的形成:原肠胚背中线的外胚层细胞向内凹陷,形成神经板,其两同时翘起形成一对纵褶,叫神经褶。神经褶又相向生长,在神经板背侧愈合;神经板 两侧亦向上包围生长;彼此接合。于是在背面出现一条中空的管道,即神经管。
脊索与中胚层的发生:在神经管形成的同时,原肠腔的背面、神经板下方的内胚层细胞向背方突出,逐渐脱离内胚层形成一条纵贯的脊索;在脊形成的同时,脊索突 起的两侧也发生若干成对的分节状突起,最后脱离内胚层,自成一腔囊,叫体腔囊,也以是中胚层。将来发育成为体腔。内胚层除了分离成脊索和中胚层外,其他部分则形成肠管的内壁。
因具有纵贯体躯一直达到前端的脊索而得名;头无明显分化,与体躯不能截然分开,故又名无头类,通称文昌鱼、矛形鱼。头索动物终生具有纵贯背部,起支撑作用的脊索;有背神经管和咽鳃裂。这些在高等脊索动物中只存在于胚胎或幼虫期,成体消失或分化为更高级的器官。
外形特征
体形似小鱼,半透明,无头和躯干之分,左右侧扁,两端较尖,故又名Amphioxus。喜栖水质清澈的浅海海底泥沙中,平时少活动,大半身体埋于泥沙中,前端露出沙外。身体除口以外,还有两个孔与外界相通,即腹孔(atriopore)或围鳃腔孔和肛门(anus)。
皮肤
皮肤有表皮(epidermis)和真皮(dermis)的分化。表皮仅由单层柱状上皮细胞组成,表皮外覆有一层角皮层(cuticle)。真皮是一薄层胶冻状结缔组织。腹面前部两侧有由皮肤下垂形成的成对的腹褶(metapleural fold). fold)。
骨骼
文昌鱼无骨质的骨骼,纵贯全身并一直向前越过神经管的脊索是主要的支持结构。无偶鳍。有1背鳍(dorsal fin)和尾鳍(caudal fin)及身体腹面的臀鳍(preanal fin),口须中有类似软骨的支持物(骨骼的前体)。
肌肉
肌肉大部分集中在背部两侧,由60多对原始末分化的呈v字型的肌节组成,角顶朝前,肌节间以结缔组织的肌隔分隔。两侧肌节交错排列,使文昌鱼可在水平方向上作弯曲运动。
消化系统
简单,由前庭、口、咽、肠、肛门组成。
口:是一套特化的取食和滤食器官。身体前端有口笠和触须,触须上有感觉细胞。口笠内是前庭,前庭后方通向口。口周围是一环形的缘膜(velum),缘膜边缘向前方伸出指状突起,称轮器(wheel organ),轮器的摆动使带有食物的水流进入口中。缘膜边缘向口中央伸出缘膜触手(velar tentacle)。触手和口笠触须具有过滤的作用,挡住泥沙,并使小的食物颗粒随水流进入口中。然后进入咽部食物留在咽内,水流经鳃裂进入围鳃腔,经腹孔排出体外。
缘膜触手(velar tentacle):缘膜边缘向口中央伸出的结构,防止泥沙等进入。
轮器(wheel organ):缘膜边缘向前方伸出的指状突起,其摆动可使带有食物的水流进入口中。
咽:发达,几占体全长的1/2。其侧壁被大量的鳃裂所洞穿。咽部的背侧和腹侧具有咽上沟和内柱,两者由围咽沟相连。咽上沟有纤毛细胞,内柱中有腺细胞和纤毛细胞,食物颗粒被内柱腺细胞分泌的粘液粘成食物团,由纤毛的摆动进入咽上沟,再向后进入肠内。内柱有富集碘的功能,与脊椎动物的甲状腺有同源关系。
肠:为一直管,在其起始处向前伸出一盲囊,进入咽的右侧,称为肝盲囊(hepatic diverticulum),其内壁是腺细胞,可分泌消
化液,相当于脊椎动物的肝脏。
肛门:开口于身体左侧。
呼吸系统
气体交换在咽部以及身体表面进行。
排泄系统
没有集中的肾脏,位于咽壁背方两侧的数十对按体节排列的肾管是文昌鱼的排泄器官。每一肾管(nephridium)是一个短而弯曲的小管,一端有肾孔开口于围鳃腔,另一端以管细胞紧贴体腔,体腔的代谢废物渗透进入管细胞和肾管,再经肾孔进入围鳃腔,排出体外。
生殖系统
雌雄异体,大约26对方形生殖腺按体节排列于围鳃腔两侧,无生殖管道,成熟生殖细胞穿过生殖腺壁、体腔壁和围鳃腔壁进入围鳃腔,随水流从腹孔排出体外。体外受精。
循环系统
闭管式循环。文昌鱼的循环系统已具有脊椎动物的基本模式。血液流动方向在腹面是从后向前,在背面是从前向后。
心脏:无心脏。
动脉系统:具有搏动功能的腹大动脉和由它发出的人鳃动脉。腹大动脉发出许多成对的人鳃动脉直接进入鳃间隔,不散成毛细血管,经过气体交换后的血液在背部汇成对的背动脉根,向前将新鲜血液供给身体前端,向后合并成背大动脉,并由此发出血管到身体各个部分。
静脉系统
由身体前端回来的血液汇成1对前主静脉,身体后端回来的血液汇成1对后主静脉,它们从两侧汇人1对总主静脉又称居维叶氏管(ductus Cuvieri)。左右总主静脉汇合处为静脉窦,并通人腹大动脉。肠部返回的血液由毛细血管网汇成肠下静脉,向前进入肝盲囊后又散成毛细血管,这条血管称为肝门静脉(hepatic portal veins)。肝盲囊的毛细血管汇集成肝静脉进入静脉窦。文昌鱼的血液无色,无血细胞,无呼吸色素,氧气通过渗透进入血液。
神经系统
脊索背部中空的神经管是文昌鱼的中枢神经系统,几乎无脑和脊髓的分化,神经管前端管腔稍膨大为脑泡(cerebral vesicle)。
外形
文昌鱼的体形略似小鱼,无明显的头部,左右侧扁,半透明,可隐约见到皮下的肌节(myomere)和腹侧块状的生殖腺;身体两端尖出,故有双尖鱼(Amphioxus)之称,又因其尾形很像矛头而名海矛。一般体长约50mm,但产于美国的加州文昌鱼(Branchiostomacaliforniense)可超过100mm,是该属中已知个体最大的一种。前端的腹面为一漏斗状的口笠(oral hood),口笠内为前庭(vestibule),内壁有轮器(wheel organ),由前庭引向位于一环形缘膜(velum)中央的口。口笠和缘膜的周围分别环生触须(cirri)及缘膜触手(velar tentacle),具有保护和过滤作用,可阻挡粗砂等物随水流进入口中。整个背面沿中线有一条低矮的背鳍(dorsalfin),往后与高而绕尾的尾鳍(caudal fin)相连。此外在肛门之前还有肛前鳍(preanal fin)。无偶鳍,只在身体前部的腹面两侧各有一条由皮肤下垂形成的纵褶,称为腹褶(metapleura fold)。腹褶和肛前鳍的交界处有一腹孔(atripore),是咽鳃裂排水的总出口,故又名围鳃腔孔。
生活方式
文昌鱼喜栖浅海水质清澈的沙滩上,平时很少活动,常把身体半埋于沙中,前端露出沙外,或者左侧贴卧沙面,借水流携带矽藻等浮游生物进入口内。夜间较为活跃,凭藉体侧肌节的交错收缩,在海水中作短暂的游泳。寿命约2年8个月左右。5~7月为生殖季节,一生中可繁殖3次,其中以最后一次产卵最多。福建沿海海区曾是中国文昌鱼生息繁衍的理想场所。1932-1956年,文昌鱼在同安县一地的捕捞区面积约有22km2,每升沙样中文昌鱼的数量达357条,当时因年产量约57吨而高居世界首位。1956年,由于厦门高(崎)集(美)海堤建成后,海况(切断了环岛的海流、沙地面积大幅度减少、盐度降低、水流减缓、淤泥沉积等)发生很大变化,再加上围海造田和乱捕滥捞等原因,严重地破坏了文昌鱼的生态环境,因而造成数量急剧下降的恶果,年产量减少了40%。1970年,当地的收购量甚至降低到只有1t左右。1986年,文昌鱼在这一海域的分布区已缩小到0.5km2,而每升沙样中的数量进一步减少到3条,使原来名闻遐迩的文昌鱼产地,在20世纪半个世纪内变得完全失去了水产捕捞价值。
内部构造特征
1.皮肤:皮肤薄而半透明,由单层柱形细胞的表皮和冻胶状结缔组织的真皮两部分构成,表皮外覆有一层角皮层(cuticle)。表皮外在幼体期生有纤毛,成长后则消失殆尽。
2.骨骼:文昌鱼尚未形成骨质的骨骼,主要是以纵贯全身的脊索作为支持动物体的中轴支架。脊索外围有脊索鞘膜,并与背神经管的外膜、肌节之间的肌隔、皮下结缔组织等连续。脊索细胞呈扁盘状,其超显微结构与双壳类软体动物的肌细胞比较相似,收缩时可增加脊索的硬度。此外,在口笠触须、缘膜触手、轮器内部也都有角质物支持,奇鳍和鳃裂的鳍条(fin rays)及鳃条(gill bar)由结缔组织支持。
3.肌肉:文昌鱼背部的肌肉厚实而腹部比较单薄,与无脊椎动物周身体壁厚薄均匀的情况不同。全身主要的肌肉是60多对按节排列于体侧的"V"字形肌节(myomere),尖端朝前,肌节间被结缔组织的肌隔(myocomma)所分开。两侧的肌节互不对称,便于文昌鱼在水平方向作弯曲运动。此外,还有分布在围鳃腔腹面的横肌和口缘膜上的括约肌等,控制围鳃腔的排水及口孔的大小。
4.消化和呼吸器官:文昌鱼靠轮器和咽部纤毛的摆动,使带有食物微粒的水流经口入咽,食物被滤下留在咽内,而水则通过咽壁的鳃裂至围鳃腔,然后由腹孔排出体外。作为收集食物和呼吸场所的咽部极度扩大,几乎占据身体全长的1/2,咽腔内的构造与尾索动物相似,也具有内柱、咽上沟和围咽沟等。文昌鱼幼体的鳃裂直接开口于体表,后来形成围鳃腔,以腹孔作为咽部鳃裂的总出水口。
咽内的食物微粒被内柱细胞的分泌物粘结成团,再由纤毛运动使它从后向前流动,经围咽沟转到咽上沟,往后推送进入肠内。肠为一直管,向前伸出一个盲囊,突入咽的右侧,称为肝盲囊(hepatic diverticulum),能分泌消化液,与脊椎动物的肝脏为同源器官。食物团中的小微粒可进入肝盲囊,被肝盲囊细胞所吞噬,营细胞内消化,大微粒在肠内分解成小微粒后,也转到肝盲囊中进行细胞内消化,未消化的物质由肝盲囊重返肠中,在后肠部进行消化和吸收。肠的末端开口于身体左侧的肛门。
咽腔是文昌鱼完成呼吸作用的部位。咽壁两侧有60多对鳃裂,彼此以鳃条分开,鳃裂内壁布有纤毛上皮细胞和血管。水流进入口和咽时,藉纤毛上皮细胞的纤毛运动,通过鳃裂,并使之与血管内的血液进行气体交换,最后,水再由围鳃腔经腹孔排出体外。有人认为文昌鱼纤薄的皮肤也具有直接从水中摄取氧气的能力。
5.血液循环:文昌鱼循环系统属于闭管式,即血液完全在血管内流动,这种情形与脊椎动物基本相同。无心脏,但是具搏动能力的腹大动脉(ventral aorta),因而被称为狭心动物。由腹大动脉往两侧分出许多成对的鳃动脉(branchial arteries)进入鳃隔,鳃动脉不再分为毛细血管,它在完成气体交换的呼吸作用后,于鳃裂背部汇入2条背大动脉根。背大动脉根内含多氧血,往前流向身体前端,向后则由左、右背大动脉根合成背大动脉(dorsal aorta),再由此分出血管到身体各部。血液无色,也没有血细胞和呼吸色素,动脉中的血液通过组织间隙进入静脉。从身体前端返回的血液通过体壁静脉(parietal vein)注入一对前主静脉(anterior cardinal vein);尾的腹面有一条尾静脉(caudal vein),收集一部分身体后部回来的血液,进入肠下静脉(subintestinal vein),大部分血液则流进2条后主静脉(posterior cardinal vein)。左、右前主静脉和两条后主静脉的血液全部汇流至一对横形的总主静脉(common cardinal vein),或称居维叶氏管(ducts Cuvieri)。左、右总主静脉会合处为静脉窦(sinus venosus),然后通入腹大动脉。从肠壁返回的血液由毛细血管网集合成肠下静脉,尾静脉的部分血液也注入其中;肠下静脉前行至肝盲囊处血管又形成毛细管网,由于这条静脉的两端在肝盲囊区都形成毛细血管,因此称作肝门静脉(hepatic portal vein)。由肝门静脉的毛细血管再一次合成肝静脉(hepatic vein)并将血液汇入静脉窦内。
起源
原索动物没有留下化石,但有一个例外。在加拿大不列颠哥伦比亚的著名的布尔吉斯页岩中发现了寒武纪中期的一种动物的化石,命名为Pikaia gracilens(皮克鱼),鱼形,约5cm,具有脊索和“》”形分节肌节,与文昌鱼很相似。毫无疑问是一种脊索动物,比最早的脊椎动物化石还早几百万年。但在缺乏其他的相关化石的情况下,不大可能决定它与最早的脊椎动物的关系。1999年11月《Nature》杂志上发表了中国学者舒德干等人在5.3亿年前澄江动物群化石中发现的“华夏鳗“的文章,华夏鳗十分像现代的文昌鱼。皮克鱼比澄江动物群晚1000万年。*
地位
⑴祖先可能是原始的无头类,与无脊椎动物有共同祖先;
⑵由于适应不同生活方式而演变为两支,一支演变为原始有头类,导向脊椎动物进化之路,另一支特化为旁支,演变成头索动物的鳃口科动物;
⑶认为头索动物是当前脊椎动物的原始类群,是脊椎动物的姐妹群。在动物学上占有重要的地位。
1、本站所有文本、信息、视频文件等,仅代表本站观点或作者本人观点,请网友谨慎参考使用。
2、本站信息均为作者提供和网友推荐收集整理而来,仅供学习和研究使用。
3、对任何由于使用本站内容而引起的诉讼、纠纷,本站不承担任何责任。
4、如有侵犯你版权的,请来信(邮箱:baike52199@gmail.com)指出,核实后,本站将立即删除。




























